Kurzgefasst
Ein hoher Pflanzenartenreichtum im Wald fördert die Vernetzung zwischen Pflanzen und Insekten

Als primäre Konsumenten der pflanzlichen Biomasse in Ökosystemen leisten pflanzenfressende Insekten, darunter Käfer, Blattwanzen, Zikaden und Heuschrecken, einen wichtigen Beitrag zum Nährstoff- und Energiekreislauf und sind ein zentraler Bestandteil von Nahrungsnetzen. Zudem tragen sie erheblich zur Insektenartenvielfalt in der Landschaft bei. Um in einem Ökosystem zu überleben, sind sie auf das Vorhandensein passender Futterpflanzen angewiesen. Je nach Bandbreite geeigneter Futterpflanzen in einem Ökosystem entstehen so unterschiedlich große und unterschiedlich robuste Netzwerke, die in erheblichem Maße durch die Landnutzung beeinflusst werden können. Wissenschaftler:innen der Biodiversitäts-Exploratorien haben in einer umfangreichen Studie nun den Einfluss der Landnutzungsintensität in Wald und Grünland auf die Vernetzung zwischen Pflanzen und Insekten und die Robustheit dieser Netzwerke gegen Artenverlust untersucht. Entscheidend war in beiden Landnutzungstypen die Netzwerkgröße, also die Artenvielfalt der beteiligten Artengruppen. Eine extensive Beweidung förderte die Robustheit der Netzwerke im Grünland dabei deutlich stärker als Mahd und Düngung. Im Wald führten Kronenöffnungen und die Einbringung von Nadelholz zu größeren Netzwerken und einer höheren Robustheit. Die Ergebnisse können in Bewirtschaftungskonzepte einfließen und zum Schutz der Insektenvielfalt beitragen.
(Titelbild: Die Birkenwanze (Kleidocerys resedae) saugt bevorzugt an Birken und profitiert somit von lichten, baumartenreichen Wäldern.)
Wie beeinflusst die Landnutzungsintensität in Wald und Grünland die Struktur und Robustheit ökologischer Netzwerke zwischen Pflanzen und pflanzenfressenden Insekten? Welche strukturellen Maße in einem ökologischen Netzwerk sind entscheidend für die Robustheit?
Die Studie befasste sich mit Netzwerkstrukturen, die je nach Ökosystem und Standort unterschiedlich aufgebaut sein können. Aber wie entwickelt sich ein ökologisches Netzwerk (Abbildung 1)?
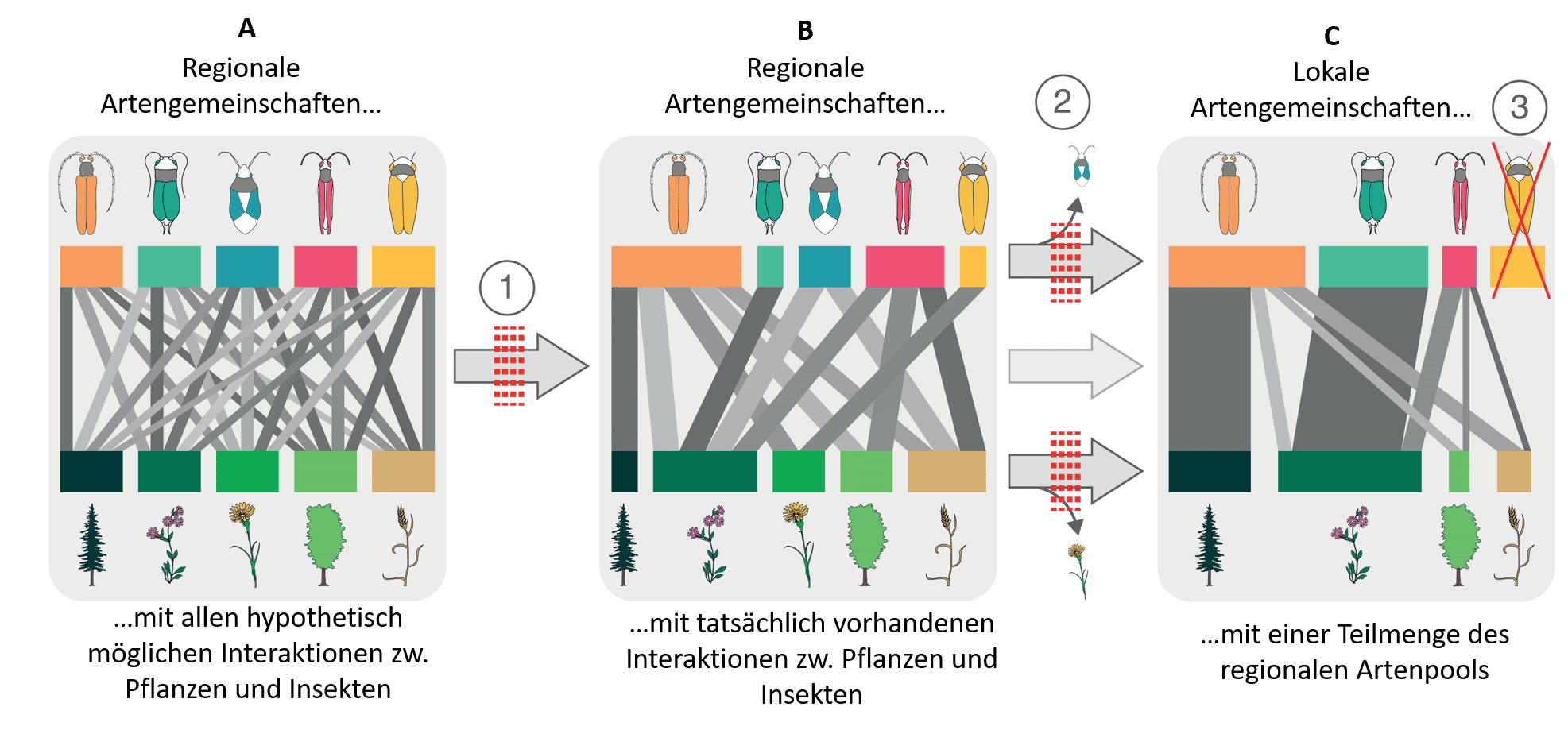
Ausgehend von einer regionalen Pflanzen- und Insektenartengemeinschaft, in der hypothetisch alle Pflanzenarten von allen Insektenarten genutzt werden können (A in Abbildung 1), sind einige Interaktionen bereits von Natur aus ausgeschlossen (1 in Abbildung 1), z. B. durch unterschiedliche Entwicklungszeiten der Pflanzen- und Insektenarten oder durch strukturelle Inkompatibilität zwischen Pflanze und Insekt. Von allen denkbaren Vernetzungen innerhalb der regionalen Artengemeinschaften können in der Natur also nur bestimmte realisiert werden (B).
Auf der lokalen Ebene, also auf einer bestimmten Wiese, Weide oder in einem Waldbestand, kommt nur eine Teilmenge der Arten der regionalen Artengemeinschaft vor (C). Hier wirken verschiedene Umwelt- und Ausbreitungsfilter, die einzelne Arten ausschließen (2). Das können Unterschiede in den Bodenbedingungen oder in der Topographie sein, aber auch Landnutzungseinflüsse. So weisen Wiesen eine andere Pflanzenartenzusammensetzung auf als Weiden, und Nadelwälder eine andere als Laubwälder. Zudem verschieben sich die Häufigkeiten von Arten in Abhängigkeit der Umweltbedingungen oder Nutzungseinflüsse. In Abbildung 1 C findet eine Insektenart (3) trotz passender Umweltbedingungen keine geeignete Futterpflanze, da ihre Wirtspflanze möglicherweise durch Mahd oder Beweidung zurückgegangen oder verschwunden ist. Die Insektenart muss daher abwandern. Ist jedoch eine Vielzahl an Futterpflanzen vorhanden (= hohe pflanzliche Artenvielfalt), ist die Wahrscheinlichkeit größer, dass Insektenarten mehrere Futterpflanzen nutzen können. Die Netzwerkgröße (Artenzahl Pflanzen x Artenzahl Insekten) macht es demnach robuster gegenüber Artenverlust.
Zudem können Netzwerke unterschiedlich organisiert sein. Man unterscheidet modulare Netzwerke, in denen Arten innerhalb von Modulen interagieren (bestimmte Insektenarten nutzen nur eine bestimmte Gruppe (z. B. Gattung) von Pflanzenarten), darüber hinaus aber wenig vernetzt sind (Abbildung 2, A) und verschachtelte Netzwerke, in denen hochspezialisierte Insekten nur eine Teilmenge an Pflanzenarten nutzen können, die weniger spezialisierte Arten nutzen (Abbildung 2, B).
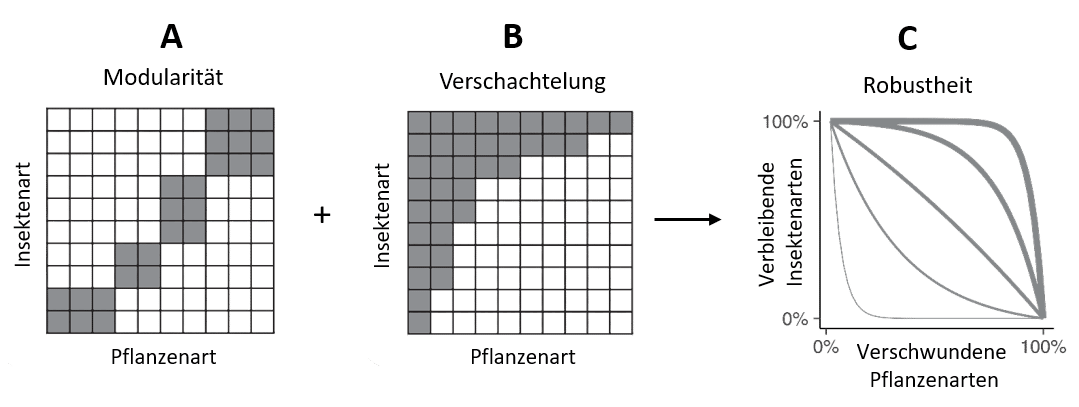
Abbildung 1B zeigt ein solches verschachteltes Netzwerk, in dem Insekt 1 alle Pflanzenarten nutzen kann, während die übrigen Arten mehr oder weniger große Teilmengen der vorhandenen Pflanzenarten nutzen. Modularität und Verschachtelung von Netzwerken werden neben der Netzwerkgröße ebenfalls als Maße zur Beschreibung der Stabilität bzw. Robustheit gegenüber Artenverlust genutzt (Abbildung 2, C). Verschachtelung wirkt i.d.R. positiv auf die Robustheit. So kann das Vorkommen einer Futterpflanze, die von vielen Insektenarten genutzt werden kann, das Risiko eines Artenverlustes abpuffern. Der Einfluss der Modularität auf die Robustheit von Netzwerken ist dagegen noch weitgehend ungeklärt. Gleiches gilt für den Einfluss der Landnutzungintensität auf die Robustheit und Strutkur von Netzwerken. Die Nutzungsgradienten der Biodiversitäts-Exploratorien (siehe Methodenpapier) können daher wichtiges Grundlagenwissen liefern.
- Zur Zusammenstellung von Pflanzen-Insekten-Netzwerken in den einzelnen Untersuchungsflächen der Biodiversitäts-Exploratorien wurden jährliche Daten von Vegetationsaufnahmen (auf 4 x 4 m-Flächen im Grünland, 20 x 20 m-Flächen im Wald) genutzt. Deckungsgrade der einzelnen Pflanzenarten wurden über die Aufnahmejahre gemittelt. Für die Baumartenzusammensetzung wurden im Wald zusätzlich die Waldinventuren der insgesamt 1 ha-großen Untersuchungsflächen herangezogen.
- Insektenarten wurden im Grünland durch Kescherfänge sowohl im Frühjahr als auch im Sommer jährlich von 2008 bis 2017 auf insgesamt 150 Flächen erfasst. Im Wald wurden Kreuzfensterfallen in ca. 2 m Höhe eingesetzt, die in den Jahren 2008, 2011 und 2014 über die ganze Vegetationsperiode (März bis Oktober) auf insgesamt 140 Flächen beprobt wurden. (siehe „Kurzgefasst“ Seibold et al. 2019). Hier wurden 139 Flächen berücksichtigt.
- Die Studie konzentrierte sich auf Käfer (Coleoptera), Wanzen (Heteroptera), Zikaden (Auchenorrhyncha, überwiegend nur Grünland) und Heuschrecken (Orthoptera, überwiegend nur Grünland). Diese Gruppen deckten im Grünland 61% der insgesamt gefangenen pflanzenfressenden (herbivoren) Arten ab, im Wald waren es 33%. Im Wald stellten auch Fransenflügler (Thysanoptera), Blattläuse (Sternorrhyncha) und Larven einen relativ hohen Anteil an Herbivoren, die schwer zu bestimmen waren und denen keine Futterpflanzen zugeordnet werden konnten.
- Um jährliche Fluktuationen bei den Insektenarten auszugleichen, wurden die Fänge pro Fläche über die Erfassungsjahre gepoolt.
- Um Netzwerke zwischen Pflanzen- und Insektenarten zu konstruieren, wurde zunächst eine Datenbank erstellt, die mögliche Interaktionen zwischen Pflanzen und Insekten auf Basis einer Literaturrecherche enthielt. Dazu wurden für alle in der Studie vorkommenden Insektenarten Information zu Futterpflanzen auf der kleinstmöglichen taxonomischen Ebene eingeholt. Unter den Baumarten konnten z. B. Kiefer und Eiche im Vergleich zu anderen Baumarten von einer Vielzahl an Insektenarten genutzt werden, darunter waren ca. 30% Spezialisten.
- Anschließend wurde für jede Untersuchungsfläche ein Netzwerk zwischen vorkommenden Pflanzen- und Insektenarten aufgestellt, basierend auf den bekannten Interaktionen in der Datenbank. Die Interaktion zwischen Insektenart und möglichen Futterpflanzen wurde als entsprechend stärker angesehen, je höher der Pflanzendeckungsgrad war.
- Für alle flächenbasierten Netzwerke wurden nun die Maße Modularität, Verschachtelung und Robustheit mit Hilfe statistischer Programme ermittelt. Daneben wurden auch die Pflanzenartenvielfalt, die Vielfalt pflanzenfressender Insekten und die Netzwerkgröße (Anzahl Pflanzenarten x Anzahl pflanzenfressender Insekten) bestimmt.
- Zunächst wurde mittels Regressionsanalysen geprüft, welchen Einfluss die Organisation von Netzwerken (Netzwerkgröße, Modularität, Verschachtelung) auf die Robustheit hat. Mit Hilfe von bayesanischen hierarchischen Modellen wurde anschließend der Einfluss der Landnutzungsintensität auf die unterschiedlichen Netzwerkstrukturen und –maße untersucht.
- Als Maß für die Landnutzungsintensität im Grünland wurde der sog. LUI-Index (Land-Use-Intensity-Index) berechnet. Dieser basiert auf jährlichen Informationen zur Stickstoffdüngung, Beweidung und Mahd. Extensiv genutztes Grünland sind ungedüngte und ungemähte Schafweiden. Eine mittlere Intensität ergibt sich für ungedüngte, einschürige Wiesen oder für Weiden mit vier Rindern/ha für ca. 50 Tage. Die höchste lokale Nutzungsintensität weisen zwei- bis dreischürige (maximal vierschürige) Wiesen auf, Weiden mit fünf bis zehn Rindern/ha für 100 bis 150 Tage oder Mähweiden, oftmals mit gleichzeitiger Düngung von bis zu 400 kg N/ha in manchen Jahren. Mahd und Düngung sind i.d.R. positiv korreliert, während Beweidung sowohl mit Mahd als auch mit Düngung negativ korreliert war.
- Im Wald ergab sich ein Bewirtschaftungsgradient durch die Berücksichtigung aktuell nicht mehr bewirtschafteter Buchenwälder, bewirtschafteter Buchen- und Buchenmischwälder und bewirtschafteter Nadelwälder (hauptsächlich Kiefer und Fichte). Ein kontinuierlicher Landnutzungsgradient analog zum Grünland wurde durch die Berücksichtigung vorangegangener Ernten, dem Anteil (z. T. standortsfremder) Nadelbaumarten und der Herkunft des Totholzes (Totholz mit Sägespuren im Verhältnis zum Gesamttotholz) errechnet (= ForMI-Index; Forest Management Index). Für unbewirtschaftete Buchenwälder wurde ein ForMI von 0.23 ermittelt, während Fichtenreinbestände den höchsten Wert mit 1.88 aufwiesen.
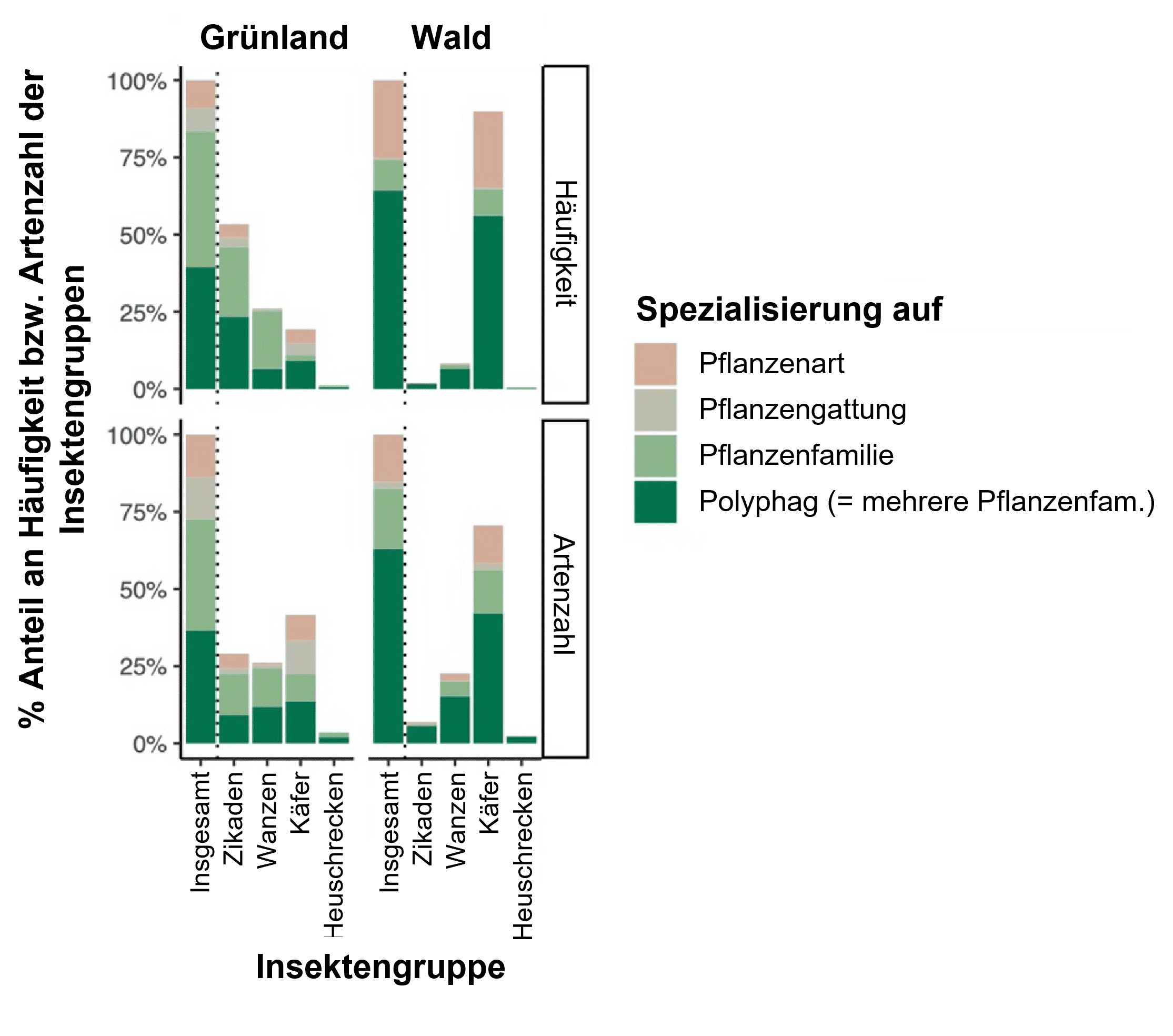
- Grünlandnetzwerke wurden auf Basis von 773 Insektenarten und 352 Pflanzenarten aufgebaut, Netzwerke im Wald bestanden aus 552 Insektenarten und 326 Gefäßpflanzenarten. Damit war die Netzwerkgröße im Grünland deutlich größer.
- Im Wald trat aber ein größerer Anteil polyphager Insektenarten auf, so dass die Verschachtelung im Wald größer war als im Grünland (Abbildung 3).
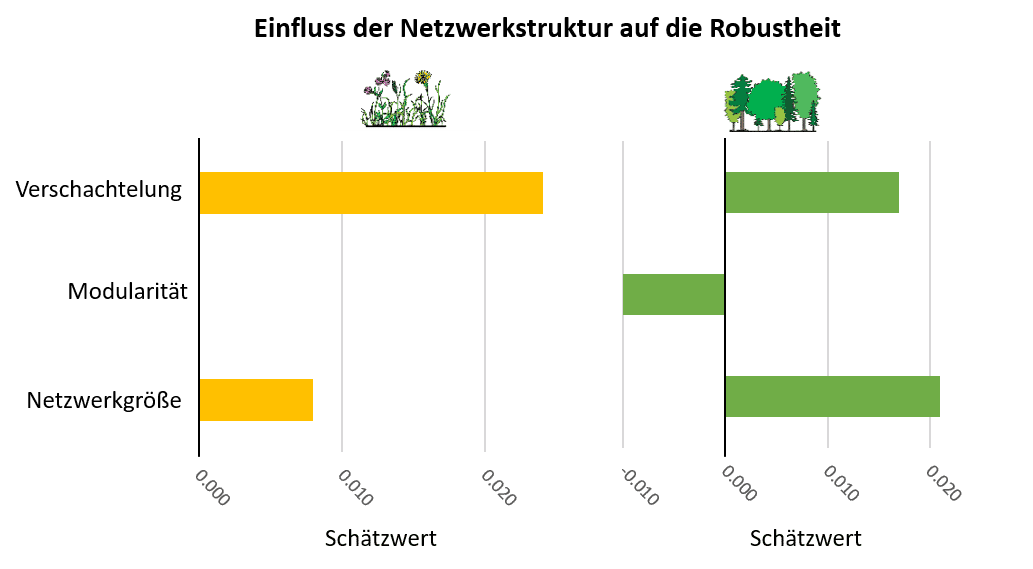
- Vor allem im Wald (aber auch im Grünland) beeinflusst die Netzwerkgröße, also die Artenzahl, die Robustheit der Netzwerke positiv (Abbildung 4). Dies unterstreicht die Bedeutung von artenreichen Systemen für die Aufrechterhaltung von Artengemeinschaften und ihrer Funktionen. Verschachtelung erhöhte die Robustheit vor allem im Grünland (aber auch im Wald) und scheint demnach ebenfalls stabilisierend zu wirken. Modularität verringerte die Robustheit im Wald leicht. Dies deutet daraufhin, dass es im Wald einige spezialisierte Beziehungen zwischen Pflanzen- und Insektenarten gibt, und der Verlust seltener Futterpflanzen zum Verlust entsprechender Insektenarten führt.
- Im Grünland war eine geringere Landnutzungsintensität (= extensive Beweidung) mit einer höheren Pflanzenartenvielfalt, Insektenartenvielfalt, Netzwerkgröße und Modularität verbunden, wobei sich dies nicht direkt auf die Robustheit im Vergleich zu einer mittleren Landnutzungsintensität auswirkte (Abbildung 5), jedoch indirekt über die Netzwerkgröße (Abbildung 4). Die Verschachtelung wurde nicht durch die Landnutzungsintensität beeinflusst. Sie wird überwiegend durch die Untersuchungsregionen mit unterschiedlichen Artenpools bestimmt.
- Aufgeteilt in die einzelnen Komponenten, wirkte eine hohe Beweidungsintensität positiv auf die Robustheit, jedoch negativ auf die Modularität (Abbildung 5). Vermutlich sorgt die häufige Störung durch das Weidevieh dafür, dass intensiv genutzte Weiden weniger durch auf bestimmte Pflanzengruppen (z. B. Gräser) spezialisierte Insekten geprägt sind, sondern durch Generalisten, die Pflanzenarten unterschiedlicher Kleinstandorte nutzen können. Auch diese Netzwerke sind dann robust.
- Intensive Mahd reduzierte sowohl die Artenvielfalt beider Gruppen als auch die Netzwerkgröße, jedoch ohne direkten Effekt auf die Robustheit. Dies deutet daraufhin, dass vor allem Spezialisten durch Mahd verloren gehen, Generalisten die Netzwerke aber aufrechterhalten und keine weiteren Artenverluste erfolgen.
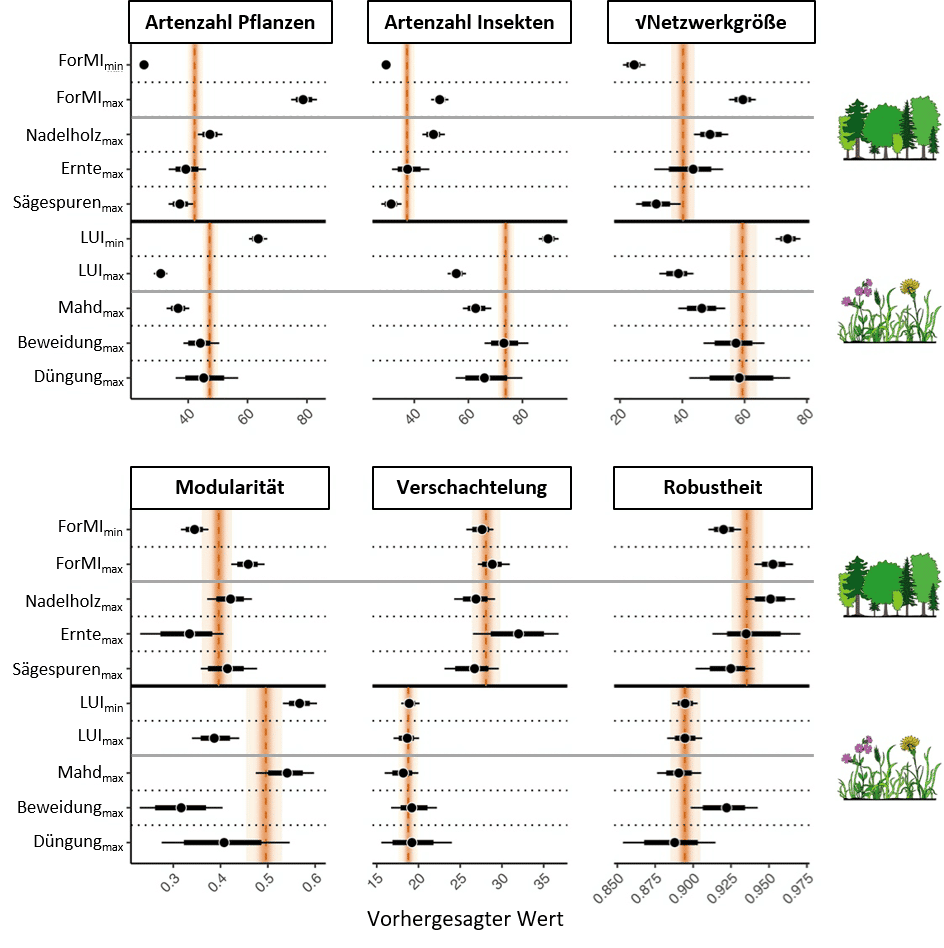
- Im Wald führte ein hoher ForMI-Index zu einer höheren Robustheit gegenüber Artenverlust im Vergleich zu geringer oder mittlerer Nutzungsintensität (Abbildung 5). Dieser Effekt ergab sich vorrangig durch die positive Wirkung der Waldbewirtschaftung auf die Pflanzenartenvielfalt, gefolgt von der Insektenartenvielfalt, was folglich die Netzwerkgröße erhöhte. Getrieben wurde dieser positive Effekt vor allem durch den Anteil an Nadelholz. Die Verschachtelung, die nach Abbildung 4 die Robustheit ebenfalls positiv beeinflusst, wurde durch die Erntemenge getrieben. Beimischung von (Nadel-)Baumarten, Bodenverwundung und Auflichtung schaffen demnach verschiedene Nischen für pflanzenfressende Insekten, vergrößern so die Netzwerke und ihre internen Beziehungen und erhöhen die Robustheit gegenüber Artenverlust.
- Sehr kleine Netzwerke zeigten sich vor allem in seit wenigen Jahrzehnten unbewirtschafteten Wäldern, die sich aktuell überwiegend in der Optimalphase befinden und allein durch die Buche dominiert sind. Ein dichtes Kronendach verringert auch die Bodenvegetation und ihre Artenvielfalt.
- Insgesamt ist die Stabilität von Pflanzen-Insekten-Netzwerken im Grünland geringer als im Wald, was sich auf häufigere Störungen und eine stärkere Spezialisierung der Arten im Grünland (Abbildung 3) zurückführen lässt.
- Wald- und Grünlandbewirtschaftung beeinflussen ökologische Netzwerke und damit verschiedene Ökosystemprozesse und –funktionen.
- Sowohl im Wald als auch im Grünland waren große Netzwerke stabiler als kleine Netzwerke. Entscheidend ist hier die Pflanzenartenvielfalt, die die Vielfalt pflanzenfressender Insekten treibt. Viele Pflanzenarten erhöhen die Chance für eine Insektenart, eine geeignete Futterpflanze zu finden und verhindern so das Abwandern dieser Art.
- Eine Landnutzung, die die pflanzliche Artenvielfalt erhöht, kann demnach zur Stabilität der Vernetzung zwischen Pflanzen- und Insektenarten beitragen und somit auch zum Erhalt der Insektenartenvielfalt.
- Im Wald kann dies durch die Beimischung verschiedener Baumarten (insbesondere Nadelbaumarten aber auch Eiche) in durch die Buche geprägte Waldökosysteme erreicht werden, und durch Kronenöffnungen (durch natürliche Störungen oder Baumernte), die die Lichtverfügbarkeit erhöhen und so die Bodenvegetation fördern. Insgesamt förderte im Wald die Nutzungsintensität die Vernetzung zwischen Pflanzen- und Insektenarten. In unbewirtschafteten Wäldern werden natürliche Absterbeprozesse in Zukunft vermutlich zu größeren Netzwerken führen.
- Im Grünland ist extensive Beweidung ökologisch günstiger als Mahd, die oft in Verbindung mit Düngung erfolgt, da sie den Pflanzen- und Insektenartenreichtum fördert. Insgesamt ist im Grünland weniger (Mahd, Weidevieh, Dünger) mehr.
- Die Ergebnisse gelten im Rahmen der in den Biodiversitäts-Exploratorien untersuchten Landnutzungsgradienten (siehe Methodenpapier), die repräsentativ sind für Mitteleuropa. Höhere Beweidungs- oder Mahdintensitäten im Grünland, sowie Kahlschläge im Wald werden nicht abgebildet. Auch echte Primärwälder sind nicht Teil der Untersuchung.
- Die Kombination von jährlichen Artenerfassungen mit Informationen aus der Literatur zu Nahrungspräferenzen der Insekten erlaubte wichtige Rückschlüsse auf die Beziehung zwischen Pflanzen und Insekten im Wald und im Grünland. Die anhand der Literatur identifizierten Interaktionen wurden jedoch nicht durch Beobachtungen im Gelände bestätigt. So können unterschiedliche Umweltbedingungen möglicherweise auch die Nahrungspräferenzen innerhalb einer Art beeinflussen.
- Im Wald wurden nur die Insektenarten in 2 m Höhe berücksichtigt. Die Artengemeinschaft im Kronenraum kann davon abweichen. Ergebnisse aus einem Jahr, im dem auch die Artengemeinschaften in der Krone erfasst wurden, bestätigen aber die Ergebnisse.
- Die Untersuchungsflächen bilden keinen kontinuierlichen Mischungsgradienten von Buche mit Nadelholz oder anderen Baumarten ab. Ein hoher Nadelholzanteil als Teil des ForMI-Index zeigt demnach den Einfluss von Fichten- bzw. Kiefernreinbeständen. Kiefer und Fichte sind laut Literatur für deutlich mehr Insektenarten geeignete Futterpflanzen als z. B. Lärche oder Douglasie. Die Ergebnisse können demnach nicht auf andere Nadelbaumarten übertragen werden.
- In dieser Studie nicht berücksichtigt wurde der Einfluss der Landschaft auf die lokale Artenvielfalt der pflanzenfressenden Insekten (siehe „Kurzgefasst“ Le Provost et al. 2021 und Seibold et al. 2019).
Eine weitere Kurzfassung zu diesem Artikel finden Sie hier.




